
Research article / Open Access
DOI:10.31488/bjg.1000146
Long COVID, Vaccine Injury and Gut Dysbiosis
Bryan Rinde1, Guanhui Bao1 , Amelia Goudzwaard1,2, Adriana C Vidal1,2, Sabine Hazan1,2
1.ProgenaBiome, LLC, Ventura, CA, USA
2.Microbiome Research Foundation, Ventura, CA, USA
*Corresponding author: Sabine Hazan, MD, ProgenaBiome, LLC, Ventura, CA, USA, Email: drhazan@progenabiome.com
Abstract
Introduction. COVID-19 infection can trigger a chronic condition known as Long COVID, which as of today, still affects millions of U.S. adults and children and remains a major public health concern. In this study, we investigated whether Long COVID patients have a distinct gut microbiome profile when compared to healthy controls. Similarly, with the prevalence of patients reporting Long COVID symptoms upon vaccination for SARS-CoV-2, we also compared the gut microbiome profiles of these patients with those of healthy controls.Methods. Fecal samples from 31 long COVID patients, 37 COVID-19 Vaccine-Injured, and 20 healthy controls were collected at ProgenaBiome laboratories. Microbiota composition was compared using shotgun methodology and next- generation sequencing (NGS) on the Illumina NextSeq with 500/550 High-Output Kits V.2.5 for 300 cycles. Data were processed through the OneCodex bioinformatics system to determine microbiota composition and relative abundances of top genera and phyla. A Mann-Whitney U test was used to evaluate statistical significance.Results. We found that Bacteroides was the dominating microbe in both Long COVID and SARS-CoV-2 Vaccine- Injured patients. Compared to healthy controls, Long COVID patients’ gut microbiome had more Bacteroides (p<0.001) and less Bifidobacterium (p<0.001) and Faecalibacterium (p=0.002). SARS-CoV-2 vaccine-injured patients had less Bifidobacterium (p<0.001) and Faecalibacterium (p=0.002), compared to healthy controls.Conclusions. These data suggest that gut dysbiosis may be associated with Long COVID and SARS-CoV-2 Vaccine- Injury. Future research is needed to confirm these findings. However, if confirmed, Long COVID and Vaccine-Injured patients may be treated for dysbiosis to alleviate symptoms.
Keywords:SARS-CoV-2, Long COVID, Vaccine-Injured, NGS, microbiome
Introduction
Since the onset of SAR-CoV-2 in December 2019 as a glob- al pandemic, many individuals have developed chronic condi- tions after exposure to the SARS-CoV-2 virus or as a reaction to SARS-CoV-2 vaccination. According to the Household Pulse Survey, 29.8% of adults in the U.S. have had or currently have post-COVID symptoms lasting longer than 3 months [1]. Often these symptoms include chronic fatigue syndrome (ME/CFS), muscle weakness, and diarrhea [2]. Understanding why these patients experience chronic symptoms has become paramount in researchers' attempts to understand the full breadth of COVID- 19’s impact on patient wellbeing that goes well beyond the acute viral infection. As SARS-CoV-2 likely induces immune dysreg- ulation through mechanisms like cytokine storms and mitochon- drial oxidative stress, focusing on the host’s immune response would be one way to address Long COVID [3,4]. As the body’s largest housing for its immune response, studying dysregulation of the gastrointestinal tract is a logical focus for elucidating is- sues of complications after host infection [3].
Beyond SARS-CoV-2, it has been shown that many chronic inflammatory disorders such as autoimmune diseases, allergies, and metabolic syndromes, are correlated to a deteriorated sym- biosis of gut microbes in the intestine [2,5]. For SARS-CoV-2, there is a higher risk for severe infection and chronic fatigue when the virus invades patients with comorbidities like chronic obstructive pulmonary disease (COPD), which could be linked to a weak gut microbiota [6,7]. In more severe cases of SARS- CoV-2, intestinal conditions such as inflammatory bowel disease (IBD) show higher prevalence, existing in approximately 17.6% of all cases [3,8].
In successfully recovered COVID-19 patients, higher Bifido- bacterium and Faecalibacterium atiters [9]. As Bifidobacterium abun- dance is known to diminish due to certain illnesses, including viral infections like influenza, it is anticipated that a similar trend specifically linked to Bifidobacterium would be of some cause bundances have been correlat- ed with increased spike IgG for severity and chronicity of symptoms during and after SARS- CoV-2 infection [2,10]. It is, indeed, observed that COVID-19 patients experiencing hair loss have previously shown to have reduced Bifidobacterium and Faecalibacterium [11].
It has been recently shown that SARS-CoV-2 vaccinated in- dividuals also experience chronic symptoms similar to those ob- served in Long-COVID [5,12-19]. Indeed, Bhattacharjee et al., 2025, in a cross-sectional study among 42 post-SARS-CoV-2 vaccine syndrome participants and 22 healthy controls enrolled in the Yale LISTEN study, found that the SARS- CoV-2 vaccinat- ed participants had diminished immune function markers com- pared to controls [12].
To evaluate whether Long-COVID and SARS-CoV-2 Vac- cine-Injury patients share a pattern in gut microbiota changes compared to healthy controls, we measured gut microbiome abundance and composition among these groups.
Methods
Study subjects who attended a GI clinic were recruited for this study. Inclusion criteria were no symptoms reported (controls), the presence of GI symptoms and/or Long COVID/Vaccine- In- jured symptoms, and aged 18 yrs. and older. Participants were excluded if they were younger than 18 yrs. old and if symptoms were severe enough to require hospitalization. A total of (n = 88) consecutive participants who attended the GI and gave writ- ten informed consent to participate in this study were included. Fecal samples from 31 Long COVID patients, 37 COVID- 19 Vaccine-Injured, and 20 healthy controls were collected at Pro- genaBiome Laboratories.
Microbiota composition was compared using DNA quantifica- tion, purification, and normalization using shotgun methodology and next-generation sequencing (NGS) on the Illumina NextSeq with 500/550 High-Output Kits V.2.5 for 300 cycles. Data were processed through the OneCodex bioinformatics system to de- termine microbiota composition and relative abundances of top genera and phyla. A Mann-Whitney U test was used to evaluate statistical significance.
Results
Patient characteristics
The demographics of all patients from the Control (n=20), Long COVID (n=31), and Vaccine- Injured (n=37) groups are presented in Table 1. Among the controls, the average age and standard error of the mean was 46±4 years; 10 of 20 (50%) were male; 18 out of 20 (90%) self- identified as non-Hispanic White, 1 (5%) identified as Asian, and 1 as Native American (5%).
Table 1. Demographics of study participants
| Control (n=20) | Vaccine Injured (n=37) | Long COVID (n=31) | |
|---|---|---|---|
| Age (mean, SEM, range) | 46, 18, 20–87 | 51, 2.6, 16–81 | 50, 2.7, 19–85 |
| Male (n, %) | 10 (50%) | 21 (57%) | 12 (39%) |
| Female (n, %) | 10 (50%) | 16 (43%) | 19 (61%) |
| Race | # of Subjects (%) | # of Subjects (%) | # of Subjects (%) |
| White | 18 (90%) | 33 (89%) | 29 (94%) |
| Black | 0 (0%) | 1 (3%) | 0 (0%) |
| Asian | 1 (5%) | 1 (3%) | 1 (3%) |
| Hispanic | 0 (0%) | 2 (5%) | 1 (3%) |
| Native American | 1 (5%) | 0 (0%) | 0 (0%) |
| # of Comorbidities | # of Subjects (%) | # of Subjects (%) | # of Subjects (%) |
| 0 | 0 (0%) | 0 (0%) | 0 (0%) |
| 1 | 0 (0%) | 21 (57%) | 20 (65%) |
| 2 | 0 (0%) | 14 (38%) | 9 (29%) |
| 3 | 0 (0%) | 2 (5%) | 1 (3%) |
| 4 | 0 (0%) | 0 (0%) | 1 (3%) |
Among the Long COVID patients, the average age and stan- dard error of the mean was 50±2.7 years; 12 of the 31 (39%) were male; 29 out of 31 (94%) self-identified as White, 1 patient (3%) identified as Asian, and one (3%) as Hispanic. The average age and standard error of the mean, for the Vaccine-Injured group was 51±2.6 years; 21 of the 37 (57%) were male; 33 of the 37 (89%) self-identified as non-Hispanic White, 1 (3%) identified as black, 1 (3%) identified as Asian, and 2 (5%) identified as Hispanic. Both Long COVID and Vaccine-Injured patients had more than one underlying comorbidity considered risk factors, including obesity, cardiovascular conditions, and being immu- nocompromised.
Microbiome diversity and composition
NextGen Sequencing analysis revealed that, at the genus lev- el, shown in Figure 1, Bacteroides are the dominating microbe in both Long COVID and Vaccine-Injured patients. Compared to healthy controls, Long COVID patients’ gut microbiome had more Bacteroides (p<0.001) and Blautia (p=0.039), while hav- ing less Bifidobacterium (p<0.001), Faecalibacterium (p=0.002), and Collinsela (p=0.004). Vaccine-Injured patients had elevated Bacteroides (p=0.168), and less Bifidobacterium (p<0.001), Faecalibacterium (p=0.002), Collinsela (p=0.002), and Rumino- coccus (p=0.034) compared to healthy controls, Figure 1. All other species did not indicate any significant difference between patient categories.
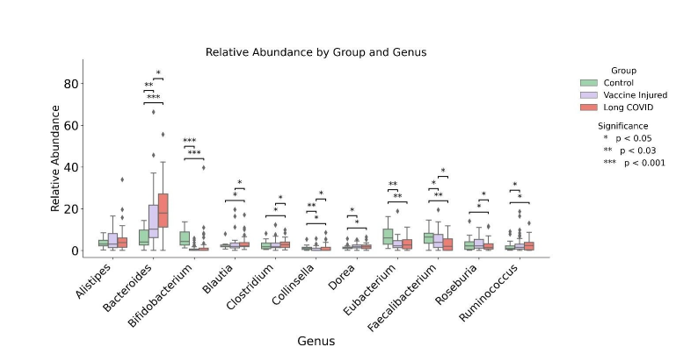
Figure 1:Controls versus Vaccine Injured and Long COVID at Genus level
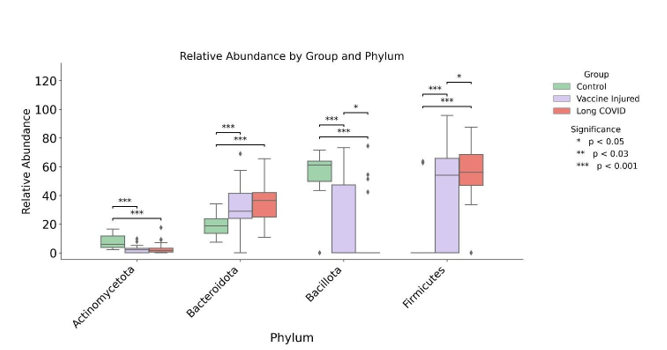
Figure 2:Controls versus Vaccine Injured and Long COVID at Phylum level
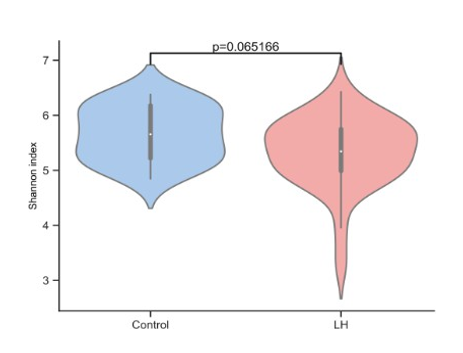
Figure 3:Gut Microbiome Diversity in Long COVID patients versus controls
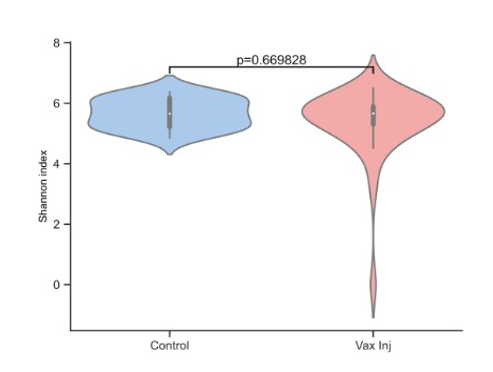
Figure 4: Gut Microbiome Diversity in Vaccine-Injured patients versus controls
At the Phylum level, Figure 2, both Long COVID and Vac- cine-Injured patients had less Actinomycetota and Bacillota, while more abundance of Bacteriodota and Firmicutes, com- pared to controls, all p < 0.05.
Long COVID patients’ gut microbiome had less overall di- versity, indicated by the Shannon Index, compared to controls, Figure 3, however, the association was not statistically signifi- cant (p=0.065). Likewise, the Shannon Index for diversity was not statistically significant when comparing Vaccine-Injured gut microbiome with controls, Figure 4.
Discussion
This study found that both Long COVID patients and SARS- CoV-2 vaccinated individuals had elevated levels of gut Bacteroi- des and decreased levels of gut Bifidobacterium and Faecalibacte- rium. If confirmed in larger studies, manipulating gut microbiota may serve as a concomitant treatment for Long COVID therapies.
Across Long COVID and post-SARS-CoV-2 vaccination syndrome cases, clinical and translational studies show charac- teristic, and often persistent, gut dysbiosis that tracks with dis- ease severity and long-term symptoms. Indeed, Gu et al. [10] in a study among 30 COVID- 19 patients, 24 H1N1 influenza, and 30 controls, reported significantly reduced bacterial diversi- ty in COVID-19 patients, enrichment of genera such as Strepto- coccus, Rothia, Veillonella, and Actinomyces, and depletion of “beneficial symbionts” while controls had higher prevalence of beneficial Bifidobacterium, Blautia, Romboutsia, and Collinsella. Using a five-taxon signature discriminated COVID-19 from con- trols with 95% confidence interval, and seven taxa distinguished COVID-19 from H1N1 also with a 95% confidence interval [10].
In a qPCR-based study of 57 patients with general, severe, or critical COVID-19 disease, Tang et al. [20] quantified large reductions in key butyrate producers across the spectrum of se- verity. For example, the median Bifidobacterium abundance fell from 5.7×106 common logarithm of copy number per gram in the general group to 4.4×106 in the severe group, and 1.4×104 in the critical group with similar stepwise declines in F. prausnit- zii, Clostridium leptum, C. butyricum, and Eubacterium rectale, while Enterococcus and Enterobacteriaceae expanded [20]. No- tably, the Ec/E ratio was able to predict mortality in critically ill patients.
Longitudinal metagenomics in smaller cohorts linked this dysbiosis to intestinal viral activity. Zuo et al. [21] followed 15 hospitalized COVID-19 patients and found that 47% of them had faecal SARS-CoV-2 by RNA metagenomics, with a “high infectivity” signature (increased 3′ - 5’ end coverage) persisting up to 6 days after respiratory PCR negativity; stools with this sig- nature were enriched in Collinsella aerofaciens, Collinsella tana- kaei, Streptococcus infantis, and Morganella morganii, whereas samples with low-to-none faecal viral activity were enriched in SCFA producers including Parabacteroides merdae, Bacteroides stercoris, Alistipes onderdonkii, and Lachnospiraceae bacterium 1_1_57FAA [21].
Larger shotgun-based cohorts extend these associations into the post-acute phase. In a prospective study of 106 patients fol- lowed for 6 months with 68 non-COVID controls, Liu et al. found that 76% met criteria for post-acute COVID-19 syndrome and at 6 months, these patients remained enriched for Rumino- coccus gnavus and Bacteroides vulgatus with depleted F. praus- nitzii and Bifidobacterium pseudocatenulatum [11].
A complementary multi-kingdom analysis of 133 patients, 296 fecal metagenomes and 79 fecal metabolomes, defined two reproducible gut configurations. One was characterized as hav- ing a low-diversity, pathogen-rich cluster associated with more severe acute disease, systemic inflammation, and a higher prev- alence of post-acute COVID-19 syndrome at 6 months, and a second cluster linked to milder illness and faster recovery [22].
Importantly, gut microbiome and SARS-CoV-2 vaccine inter- actions appear to differ in individuals with pre-existing immuno- compromised conditions. In a cohort of 143 individuals involv- ing people living with HIV and receiving BNT162b2, Ray et al. [9] showed that baseline microbiota composition and diversity predicted vaccine immunogenicity. High spike IgG and CD4+ T-cell responders exhibited lower α-diversity but relative enrich- ment of Bifidobacterium and Faecalibacterium, whereas Cloac- ibacillus was associated with poor antibody responses- suggest- ing that pre-existing microbiome configurations can modulate both the magnitude and quality of vaccine-induced immunity [9]. With evidence that specific gut bacteria influence both sus- ceptibility to and outcomes of SARS-CoV-2 infection, these findings support a model in which SARS-CoV-2 illness, Long COVID, and post-vaccination syndromes arise on a background of gut microbial dysregulation and, in turn, further perpetuate this dysbiosis.
These microbial shifts may influence immune regulation that contributes to the wide range of persistent symptoms. Possible mechanisms have been identified in earlier studies which may underly the myriad of both Long COVID and vaccine-relat- ed adverse events but have not been definitively linked to the microbiota. Marked by cytokine storms, COVID-19 infection reflects a profound immune overactivation [24]. Studies have highlighted a persistent small subset of patients that do not re- spond positively upon vaccination, sometimes reporting symp- toms within a day after vaccination [22]. In patients experiencing post-vaccination syndromes (PVS), immunological imbalances have been reported with specific signatures compared to healthy controls, including reduced circulating memory and effector T cells, and increased TNFα-producing CD8+ T cells along with an increased likelihood of Epstein-Barr virus reactivation [12]. A strong correlation was shown between the number of vaccine doses and plasma anti-S IgG. In cases of vaccine-induced im- mune thrombotic thrombocytopenia (VITT), immune responses have been shown to trigger the production of anti-platelet factor 4 (PF4) antibodies [17]. Post-vaccination autoimmune myocar- ditis has been linked to CD4+ T cell– mediation, with elevated levels of IL-1q and IL-1ra, cytokines central to the vaccine im- mune response, especially for male patients that suggests a sex hormone influence [18].
Neurological complications, including Guillain-Barré syn- drome for patients with a predisposition, have been observed, likely due to the production of spike proteins that leads to au- toimmune demyelination [17]. Moreover, free circulating spike protein has been detected exclusively in symptomatic individu- als and has been correlated with cardiac troponin T, indicating a possible role in cardiac inflammation and injury [18].
The vast diversity of bacteria in the human body suggests a range of natural mechanisms in immune regulation, many of which remain poorly understood [25]. Some bacteria actively participate in the body’s regulation of the immune system, acting through a pro- inflammatory means while others work through an anti-inflammatory mechanism [8]. As key components of im- mune regulation depend on microbial metabolites, disruptions in the microbiome may influence both susceptibility to infection and disease severity [5].
As the primary target for active SARS-CoV-2, angioten- sin-converting enzyme 2 (ACE2) receptors indicate likely the first effected cellular mechanism and the first connection the gut microbiome has to bodily infection. The intestine is known to express a higher density of ACE2 receptors than most other tis- sue [19,22]. While ACE2 allows the entry of SARS-CoV-2 into the body through the intestine, it allows barrier function impair- ment, including nutrient absorption, while acting as a regulator for the gut microbiota [10,13,14].
Short-chain fatty acids (SCFAs) must also be considered. SC- FAs are utilized in the production of O-glycans that aid in the permeability of the intestinal wall for translocation of pathogens and toxins [14,26]. The permeability of the intestinal wall is af- fected by SCFAs, L- isoleucine, and bile acids. Losing the pro- duction of SCFAs through loss of Bifidobacterium, Ruminicoc- cus, and Clostridium also elevate the inflammatory response by disrupting the adaptive immune system by not regulating T-cell development through GPCR. With a reduction in SCFAs, as well as L-isoleucine, the largest immune reservoir housed in the intestine is weakened [27]. Patients with chronic inflammatory symptoms will also express higher levels of pro-inflammatory cytokines IL-18, IL-1 beta, and IL-6, which increase lactic acid production that can lead to further pain [28,29].
Correlated to gut microbiota health, higher IgG titers in some individuals were associated with greater abundance of Bifido- bacterium and Faecalibacterium, though paradoxically accom- panied by reduced alpha diversity [9]. Along with Faecalibac- terium prausnitzii, Roseburia and Ruminoccus, are butyrate producing bacteria whose reduced abundances have also been correlated with negative outcomes of SARS-CoV-2 infection, including hair loss [11].
While most changes in the microbiome are decreases in bac- terial abundances, the biggest change appears in the increase of Bacteroides, which have been shown to exacerbate pro-in- flammatory responses in the intestine through the release of TNF- α and immune cells such as IFN-γ regulatory CD4+ T cells [5,30,31]. A correlation is seen in the overabundance of Bacteroides with the overexpression of lectin reg III beta fusion protein which is already known to be involved in most states of inflammation in the intestine [32,33].
With the goal of treating conditions such as inflammatory bowel disease through modulation of the gut microbiome, in- terventions including probiotic supplementation and fecal mi- crobiota transplantation (FMT) have shown therapeutic benefit. These approaches have been associated with increased relative abundances of key beneficial taxa, particularly
Bifidobacterium and Faecalibacterium [34,35]. If severity of SARS-CoV-2 infection does present a correlative linkage to the gut microbiome, similar treatments would be beneficial for patients experiencing chronic symptoms post-infection or post-vaccination.
Study limitations include the relatively small sample size, and analyses were not adjusted for confounders. However, the observed gut microbiota alterations were distinguishable across patient groups.
Conclusions
While acute SARS-CoV-2 infection was initially the concern, attention has shifted toward the growing number of individuals experiencing long-term effects following infection or vaccina- tion. Cases of Long COVID and adverse reactions to SARS- CoV-2 vaccines can persist for years, with symptoms including, but not limited to, muscle pain, fatigue, shortness of breath, and anxiety. Many individuals are forced to adjust their lifestyles to accommodate the ongoing nature of chronic illness, often with- out knowing when or if symptoms will subside. This uncertainty highlights an urgent need for effective and lasting therapeutic strategies. In an effort to explore the role of microbial dysbio- sis in these persistent conditions, our findings reveal consistent alterations in the gut microbiota, which are common among af- fected individuals.
With targeted interventions such as probiotic treatment, mod- ulation of the gut microbiome may offer a gradual path to recov- ery and an adjunctive approach to Long COVID and vaccine injury management.
Declarations
Ethics approval and consent to participate
This study was approved by the institutional review board of IRCM IRB. Consent to participate in this study from each patient was agreed via signed informed consent.
Consent for publication
Agreed to by patients on the signed informed consent, demon- strating that the patient understood the procedures and the pur- pose of the study.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Competing interests
Dr. Hazan is the CEO of Progenabiome and the owner of Mi- crobiome Research Foundation.
Funding
This study was funded by ProgenaBiome.
Authors’ contributions
Conceptualization, BR and SH; Methodology, BR, AG, and SH; Investigation, AV and SH. Writing-original draft, BR; Writ- ing-review and editing, BR, AV, and SH Supervision, SH; Fund- ing acquisition, SH. All authors read and approved he final man- uscript.
References
1. U.S. Census Bureau. Household Pulse Survey: Phase 4.2 [Internet]. U.S. Department of Commerce; [cited 2024 Oct 22]. Available from: https://www.census.gov/data-tools/demo/hhp/#/?measures=LONGCOVID_1
2. Zuo T, Wu X, Wen W, Lan P. Gut microbiome alterations in COVID-19. Genomics Proteomics Bioinformatics. 2021 Oct;19(5):679–688.
3. Chaves Andrade M, Souza De Faria R, Avelino Mota Nobre S. COVID-19: Can the symptomatic SARS-CoV-2 infection affect the homeostasis of the gut-brain-microbiota axis? Med Hypotheses. 2020 Nov;144:110206.
4. Georgieva E, Ananiev J, Yovchev Y, Arabadzhiev G, Abrashev H, Abrasheva D, et al. COVID-19 complications: Oxidative stress, inflammation, and mitochondrial and endothelial dysfunction. Int J Mol Sci. 2023 Oct 4;24(19):14876.
5. Belkaid Y, Hand TW. Role of the microbiota in immunity and inflammation. Cell. 2014 Mar;157(1):121–141.
6. Bouazza B, Hadj-Said D, Pescatore KA, Chahed R. Are patients with asthma and chronic obstructive pulmonary disease preferred targets of COVID-19? Tuberc Respir Dis. 2021 Jan;84(1):22–34.
7. Vu QM, Fitzpatrick AL, Cope JR, Bertolli J, Sotoodehnia N, West TE, et al. Estimates of incidence and predictors of fatiguing illness after SARS-CoV-2 infection. Emerg Infect Dis. 2024 Mar;30(3):539–547.
8. Alharbi KS, Singh Y, Hassan Almalki W, Rawat S, Afzal O, Alfawaz Altamimi AS, et al. Gut microbiota disruption in COVID-19 or post-COVID illness: Association with severity biomarkers and possible role of pre/probiotics. Chem Biol Interact. 2022 May;358:109898.
9. Ray S, Narayanan A, Vesterbacka J, Blennow O, Chen P, Gao Y, et al. Impact of the gut microbiome on immunological responses to COVID-19 vaccination. NPJ Biofilms Microbiomes. 2023 Dec 20;9(1):104.
10. Gu S, Chen Y, Wu Z, Chen Y, Gao H, Lv L, et al. Alterations of the gut microbiota in patients with coronavirus disease 2019 or H1N1 influenza. Clin Infect Dis. 2020 Dec 17;71(10):2669–2678.
11. Liu Q, Mak JWY, Su Q, Yeoh YK, Lui GCY, Ng SSS, et al. Gut microbiota dynamics in patients with post-acute COVID-19 syndrome. Gut. 2022 Mar;71(3):544–552.
12. Bhattacharjee B, Lu P, Monteiro VS, Tabachnikova A, Wang K, Hooper WB, et al. Immunological and antigenic signatures associated with chronic illnesses after COVID-19 vaccination [Internet]. 2025. Available from: https://www.medrxiv.org/content/10.1101/2025.02.18.25322379v1
13. Camilleri M. Leaky gut: Mechanisms, measurement and clinical implications in humans. Gut. 2019 Aug;68(8):1516–1526.
14. Verdecchia P, Cavallini C, Spanevello A, Angeli F. The pivotal link between ACE2 deficiency and SARS-CoV-2 infection. Eur J Intern Med. 2020 Jun;76:14–20.
15. Faksova K, Walsh D, Jiang Y, Griffin J, Phillips A, Gentile A, et al. COVID-19 vaccines and adverse events of special interest. Vaccine. 2024 Apr;42(9):2200–2211.
16. Schultz NH, Sørvoll IH, Michelsen AE, Munthe LA, Lund-Johansen F, Ahlen MT, et al. Thrombosis and thrombocytopenia after ChAdOx1 nCoV-19 vaccination. N Engl J Med. 2021 Jun 3;384(22):2124–2130.
17. Lamprinou M, Sachinidis A, Stamoula E, Vavilis T, Papazisis G. COVID-19 vaccines adverse events: Potential molecular mechanisms. Immunol Res. 2023 Jun;71(3):356–372.
18. Buoninfante A, Andeweg A, Genov G, Cavaleri M. Myocarditis associated with COVID-19 vaccination. NPJ Vaccines. 2024 Jun 28;9(1):122.
19. Du M, Cai G, Chen F, Christiani DC, Zhang Z, Wang M. Multiomics evaluation of gastrointestinal characteristics of COVID-19. Gastroenterology. 2020 Jun;158(8):2298–2301.e7.
20. Tang L, Gu S, Gong Y, Li B, Lu H, Li Q, et al. Correlation between intestinal bacteria and COVID-19 severity. Engineering. 2020 Oct;6(10):1178–1184.
21. Zuo T, Liu Q, Zhang F, Lui GCY, Tso EY, Yeoh YK, et al. SARS-CoV-2 faecal activity and gut microbiota composition. Gut. 2021 Feb;70(2):276–284.
22. Liu Q, Su Q, Zhang F, Tun HM, Mak JWY, Lui GCY, et al. Multi-kingdom gut microbiota analyses define COVID-19 severity. Nat Commun. 2022 Nov 10;13(1):6806.
23. Alharbi KS, Singh Y, Hassan Almalki W, Rawat S, Afzal O, Alfawaz Altamimi AS, et al. Gut microbiota disruption in COVID-19 or post-COVID illness. Chem Biol Interact. 2022 May;358:109898.
24. Klawitter F, Ehler J, Bajorat R, Patejdl R. Mitochondrial dysfunction in ICU-acquired weakness. Int J Mol Sci. 2023 Mar 14;24(6):5516.
25. Gavzy SJ, Kensiski A, Lee ZL, Mongodin EF, Ma B, Bromberg JS. Bifidobacterium mechanisms of immune modulation. Gut Microbes. 2023 Dec 18;15(2):2291164.
26. Brown JA, Sanidad KZ, Lucotti S, Lieber CM, Cox RM, Ananthanarayanan A, et al. Gut microbiota metabolites protect against SARS-CoV-2. Gut Microbes. 2022 Dec 31;14(1):2105609.
27. Yeoh YK, Zuo T, Lui GCY, Zhang F, Liu Q, Li AY, et al. Gut microbiota reflects COVID-19 severity. Gut. 2021 Apr;70(4):698–706.
28. Chen G, Wu D, Guo W, Cao Y, Huang D, Wang H, et al. Clinical and immunological features of COVID-19. J Clin Invest. 2020 Apr;130(5):2620–2629.
29. Lu J, Wei Z, Jiang H, Cheng L, Chen Q, Chen M, et al. Lactate dehydrogenase and mortality in sepsis. J Surg Res. 2018 Aug;228:314–321.
30. Schirmer M, Smeekens SP, Vlamakis H, Jaeger M, Oosting M, Franzosa EA, et al. Linking gut microbiome to cytokine production. Cell. 2016 Nov;167(4):1125–1136.e8.
31. Schirmer M, Kumar V, Netea MG, Xavier RJ. Variation in human cytokine production. Curr Opin Immunol. 2018 Oct;54:50–58.
32. Miki T, Goto R, Fujimoto M, Okada N, Hardt WD. RegIIIβ prolongs gut colonization in Salmonella. Cell Host Microbe. 2017 Feb;21(2):195–207.
33. Stelter C, Käppeli R, König C, Krah A, Hardt WD, Stecher B, et al. RegIIIβ kills competing gut microbiota. PLoS One. 2011 Jun 9;6(6):e20749.
34. O’Callaghan A, Van Sinderen D. Bifidobacteria in human gut microbiota. Front Microbiol. 2016 Jun 15;7:925.
35. Zhou B, Pang X, Wu J, Liu T, Wang B, Cao H. Gut microbiota in COVID-19. Gut Microbes. 2023 Dec 31;15(1):2201157.
Received: February 06, 2026;
Accepted: February 21, 2026;
Published: March 05, 2026.
To cite this article : Rinde B, Bao G, Goudzwaard A, Vidal AC, Hazan S. Randomized single-blind clinical trial com- paring left lateral decubitus position vs. supine position for colonoscopies: A multicenter study. Br J Gastroenterol. 2026; 8(1): 326-332. doi: 10.31488/bjg.1000146.
© The Author(s) 2026. This is an open access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/).